
Ремарка
Эта тема входит в цикл "Аквариумистика по‑научному".
Оглавление цикла: здесь
Оглавление цикла: здесь
В своё время было сломано очень много копий в среде аквариумистов по поводу фотосинтеза. И не самого этого процесса, а именно про его темновую фазу.
Почему-то все (ну почти) считают, что если фаза называется "темновой", то и происходит она ночью. То есть тогда, когда свет на растения не попадает.
Сегодня я хочу немного прояснить этот вопрос.
Сразу скажу, будет много не всем понятных научных терминов, но я попытаюсь насколько смогу разъяснить и термины и процессы.
Ну и начнём с теории.
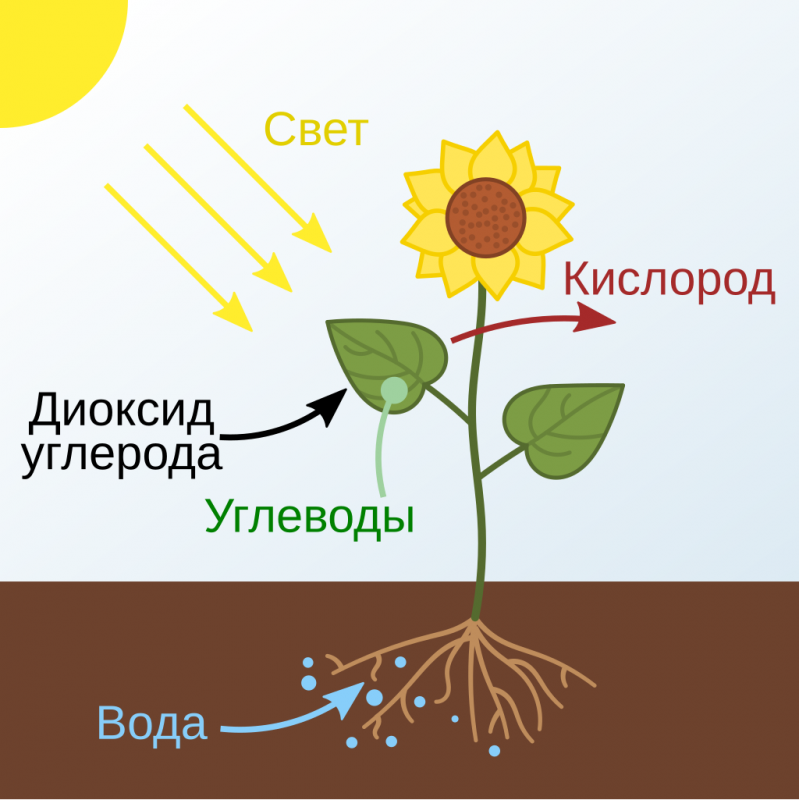
Реакции фотосинтеза традиционно делят на две стадии - темновую и световую. И тут важно понимать, что, несмотря на название, обе стадии идут только на свету! Это объясняется тем, что многие ферменты, катализирующие ключевые реакции, активны только на свету. Более того, для многих из них обнаружена зависимость от циркадных ритмов, внутренних часов организма, определяющих суточную ритмику различных процессов.Википедия писал(а):Фотосинтез (от др.-греч. φῶς - "свет" и σύνθεσις - "соединение", "складывание", "связывание", "синтез") - сложный химический процесс преобразования энергии видимого света (в некоторых случаях инфракрасного излучения) в энергию химических связей органических веществ при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл у бактерий и бактериородопсин у архей).
В физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция - совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндергонических реакциях, в том числе превращения углекислого газа в органические вещества.
Световая фаза фотосинтеза включает в себя разнообразные процессы, основная цель которых - преобразование солнечной энергии в энергию химических связей.
Фотосинтез - процесс с крайне сложной пространственно-временной организацией.
Разброс характерных времен различных этапов фотосинтеза составляет 19 порядков: скорость процессов поглощения квантов света и миграции энергии измеряется в фемтосекундном интервале (10−15 с), скорость электронного транспорта имеет характерные времена 10−10-10−2 с, а процессы, связанные с ростом растений, измеряются днями (105-107 с).
Также большой разброс размеров характерен для структур, обеспечивающих протекание фотосинтеза: от молекулярного уровня (10−27 м3) до уровня фитоценозов (105 м3).
В фотосинтезе можно выделить отдельные этапы, различающиеся по природе и характерным скоростям процессов:
- фотофизический;
- фотохимический;
- химический:
- реакции транспорта электронов;
- "темновые" реакции или циклы углерода при фотосинтезе.
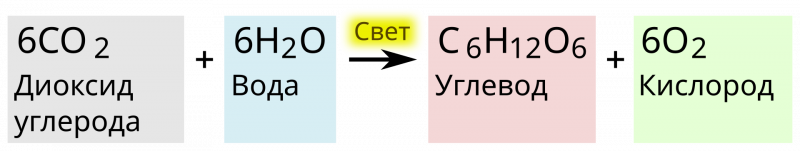
Непосредственно фиксация углекислого газа, то есть - превращение углерода из окисленной неорганической формы в восстановленную органическую, происходит в восстановительном цикле (пути) или иначе - цикле Кальвина.
Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий:
- карбоксилирования;
- восстановления;
- регенерация акцептора CO2.
Во второй стадии ФГК в два этапа восстанавливается. Сначала она фосфорилируется АТФ, затем дефосфорилируется и восстанавливается до альдегидной и образуется глицеральдегид-3-фосфат - фосфорилированный углевод (ФГА).
В третьей стадии участвуют 5 молекул ФГА, которые через образование 4-, 5-, 6- и 7-углеродных соединений объединяются в 3 5-углеродных рибулозо-1,5-бифосфата, для чего необходимы 3 АТФ. Наконец, две ФГА необходимы для синтеза глюкозы. Для образования одной её молекулы требуется 6 оборотов цикла, 6 CO2, 12 НАДФН и 18 АТФ.
То есть, цикл Кальвина представляет собой светонезависимый набор фотосинтетических реакций. Он использует аденозинтрифосфат (АТФ) и никотинамидадениндинуклеотидфосфат (НАДФН), образующиеся в ходе светозависимых реакций, для преобразования атмосферного CO2 в сложные углеводы. Цикл Кальвина также регенерирует аденозиндифосфат (АДФ) и НАДФ для светозависимой реакции.
Не запоминаем!
Если это было/будет сложно, можно пропустить! 
Пожалуй, RuBisCO - самый распространенный фермент на Земле и при этом очень древний. Содержание RuBisCO может составлять порядка 50% массы растворимых белков зеленых листьев, при этом концентрация фермента на порядки выше концентрации СО2, что абсолютно нехарактерно для ферментативных реакций.
Как было видно, реакции, протекающие при темновой фазе фотосинтеза в световой энергии не нуждаются и даже больше позволяют получить вещества, необходимые для дальнейшего протекания уже светозависимых реакций. Так проходит цикл за циклом фотосинтез.
Я думаю, вопрос о темновой фазе снят! И световая фаза и темновая проходят исключительно при наличии света. Просто, в световой фазе энергия света присутствует, а в темновой нет!
Но я хочу продолжить. Поскольку, уже упоминавшийся фермент RuBisCO играет и в аквариумистике (в выращивании водных растений) существенную роль.
Без Rubisco рост растений невозможен. Это один из самых важных участников фотосинтеза.
Rubisco отвечает непосредственно за потребление CO2, а другой энзим Rubisco activase - за активацию и регуляцию самого Rubisco. Rubisco activase помогает перейти Rubisco из неактивного состояния в активное. Это очень существенно. Только активная форма Rubisco может участвовать в процессе фотосинтеза. Например, если в клетках растения больше неактивного Rubisco, чем активного, фотосинтез замедляется.
При длительной большой подаче СО2 фиксация углерода при помощи Rubisco значительно меньше, то есть потребление CO2 зависит от этого энзима меньше. При падении концентрации CO2 снижение активности Rubisco равно фиксации СО2 при большом количестве азота, но падает на треть при недостатке азота! Это означает, что растения меньше потребляют CO2, и это замедляет их рост. При недостатке азота активность Rubisco в целом заметно ниже, и даже при большой подаче CO2 рост увеличивается совсем незначительно.
При снижении подачи CO2 не имея достаточно азота, растения просто не смогут выработать достаточно Rubisco, и фотосинтез СИЛЬНО уменьшится до малозаметного роста, а водоросли быстро адаптировавшись к измененной концентрации СО2 на фоне избытка питательных веществ, начнут быстро размножаться. Когда же азота достаточно, рост растений от большой подачи CO2 гораздо лучше, а в случае падения концентрации CO2 растения имеют в своем распоряжении достаточно азота, чтобы выработать много Rubisco и увеличить потребление малых концентраций CO2, так что рост снижается не так сильно, как при недостатке азота.
Что происходит при смене стабильно большой концентрации CO2 на его недостаток?
Об этом хорошо написал Tom Barr.
Итак, энзимы Rubisco и Rubisco activase ответственны за фиксацию СО2, а, значит, и фотосинтез.Tom Barr писал(а):Растения имеют очень мало Rubisco так как адаптировались к высоким концентрациям CO2 и теперь "ленивы". Много света требует более быстрого потребления питательных веществ. Ваши растения не могут быстро отреагировать на падение концентрации CO2. Чтобы выработать больше энзимов растениям требуется время. Им нужно несколько дней или даже недель чтобы выработать больше. Так что, если концентрация CO2 падает с 30мг/л до 15мг/л в течение дня, общий результат будет таким, что растениям некоторое время не будет хватать CO2. Это не явно выраженный эффект, существует широкая градация недостатка CO2. Через несколько недель стабильных условий растения снова будут в норме, но водоросли уже будут крепкими, чтобы было легко от них избавиться.
Когда СО2 много, Rubisco activase мало потому, что дополнительная помощь для ассимиляции СО2 не нужна. Когда же СО2 мало, растения активизируют выработку Rubisco activase который переводит из неактивного в активное состояние энзим Rubisco, что увеличивает фиксацию СО2. Чтобы это произошло Rubisco activase требует достаточного количества азота - нет азота нет и Rubisco activase, а значит и фиксации пониженных концентраций СО2.
Очень наглядно зависимость темпов роста от наличия Азота и CO2 показана в опыте Ole Pedersen с Riccia fluitans.
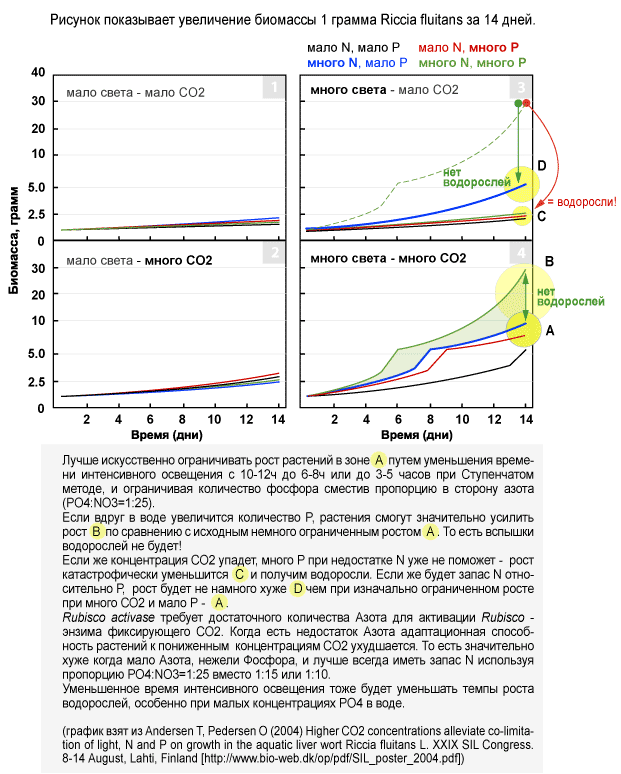
При низкой концентрации СО2 и недостатке азота рост практически останавливается (рис.3 - черная линия, зона C). Если же будет много азота рост замедлится, но не настолько, чтобы растения начали выделять питательные вещества (ПВ) и давать шансы для водорослей (3 - синяя линия, D) - азота будет достаточно для активации Rubisco и адаптации к потреблению пониженных концентраций CO2.
При достатке CO2 смена малого недостатка азота на недостаток фосфора будет проходить вообще незаметно для баланса - рост одинаковый, умеренный (рис. 4 синяя И красная линия). Зато если вдруг появится избыток фосфатов (перекормили рыб, не подменили воду пару недель или после стрижки растений…) растения будут иметь очень большой запас ускорения роста в ШЕСТЬ раз (рис.4 зеленая линия, зона B) чтобы справиться с временным повышением концентрации фосфатов. С другой стороны, при достатке CO2 и небольшом недостатке фосфора всплеска роста водорослей не будет, потому что растения продолжают довольно хорошо расти (рис. 4 синяя линия, зона А).
Наихудшая ситуация - значительное уменьшение подачи CO2 (рис.3) при интенсивном освещении. CO2 это самый НЕстабильный элемент в нашей искусственной экосистеме. Если в это время азота меньше относительно фосфора или и того и другого много(!) мы сразу же перейдем из зоны умеренного роста А в зону С, что в ЧЕТЫРЕ-ПЯТЬ раза медленнее. Если фосфора будет много как и азота при недостатке CO2, растения все равно радикально замедлят рост (C). При падении концентрации CO2 Rubisco activase требует больше азота еще больше, усугубляя его недостаток, и мы быстро скатываемся до почти полной остановки роста (с А до C в 4-5 раз, вместо D в 2 раза). Удар по системе ДВОЙНОЙ: выброс PO4 + снижение потребления и без того малых концентраций CO2 из за недостатка азота! Растения почти прекратят рост, начнут выделять избыток фосфора, и на листьях неизбежно быстро появятся водоросли.
Если же азота будет с запасом относительно фосфора (PO4:NO3=1:15-25), рост все еще будет весьма хорошим и падение концентрации CO2 переведет нас из зоны А в зону D, уменьшив рост всего лишь в ДВА раза вместо 4-6 благодаря тому, что Rubisco activase имеет достаточно азота для активации Rubisco и потребления понизившихся концентраций CO2 (рис.3 синяя линия). Это и дает СТАБИЛЬНОСТЬ. При падении концентрации CO2 смещение пропорции PO4:NO3 в сторону NO3 (с 1:7 до 1:15-25) замедляет рост растений значительно меньше. Нет азота - нет не только потребления PO4, но и CO2. Можно даже винить в стагнации растений и росте водорослей недостаток CO2, хотя его будет достаточно. Сначала нужно убедиться в достатке азота, а затем в достатке всего остального! Недостаток азота на порядок хуже, чем недостаток фосфора!
Таким образом, при снижении подачи CO2 главное, чтобы растения имели достаточно азота для усиления производства Rubisco и их рост не уменьшался сразу в несколько раз. Чтобы рост растений при достатке CO2 был не таким бурным и при падении CO2 не уменьшался в ШЕСТЬ раз, очевидно лучше иметь достаточно много азота.
Вывод. Что при малой подаче СО2, что при большой, малой интенсивности освещения или большой, ВСЕГДА следует вносить азота с запасом, особенно при малой подаче или внезапном падении CO2 (Rubisco помогающий фиксировать малые концентрации CO2 требует много АЗОТА), а соотношение PO4:NO3 в удобрении лучше сместить в сторону азота и вносить PO4:NO3 в пропорции 1:20-25. Это обезопасит от сильного замедления роста растений и возникновению прогресса в росте водорослей.
Вот такой практический вывод из теории фотосинтеза!
Спасибо за внимание.

В статье использованы материалы из интернета